Theodosius Dobzhansky: Variations on the theme of variation

"Nothing in Biology Makes Sense Except in the Light of Evolution." -- Dobzhansky (1973)
In 1927, Russian emigree and beetle taxonomist, Theodosius Dobzhansky waltzed-into Thomas Morgan's Columbia fruitfly lab. Dobzhansky's love of fruit flies became consequential in helping to answer the big question: Do we have a theory which accounts-for the level of genetic diversity observed in nature? Had it not been for him, it's an open question as to whether Darwin's biology and Mendel's laws would have been linked successfully during the Great Synthesis (1920-50). Without such linkage, evolutionary theory could not explain the spectrum of variation shown by past organisms in the fossil record and present organisms in nature.
In a landmark book written in 1937, Genetics and the Origin of Species, Dobzhansky carved-out the "new understanding" with razor-like sharpness, a comprehensiveness and a mastery of both biological theory and experimental data that no other biologist of his day could muster. A recurring theme, shot through Dobzhansky's work, was his nisus to confront patterns of observed variation.
"It follows that any difference between individuals and populations which can be expressed as a function of gene differences is to that extent due in the last analysis to mutational changes." -- Dobzhansky (1937) [1]
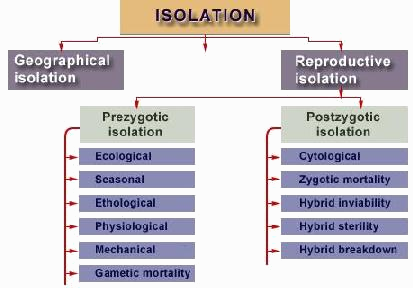
Late in the 1930s, Dobzhansky discovered "isolation mechanisms", i.e. finely-tuned sex traits, other genetic factors (and to him physical barriers in the environment) which impede gene flow between populations. [2] Without these mechanisms incorporated into evolutionary theory, it would under-explain the level of species-diversity, found in nature.
Dobzhansky was one of the first biologists to peg genetic drift as responsible for much of the variability seen among organisms and populations. By the mid-1930s, he lined-up the drivers and laws of evolution which are recognized today: natural selection, sexual selection, Mendelian inheritance and genetic drift. Eventually, Dobzhansky backed-down on the importance of drift. His (and others) demotion of drift to near-evolutionary insignificance is considered watershed in the history of evolutionary biology. S.J. Gould pronounced it "the hardening of the Synthesis". [3] A question, central to evolution, was (and remains): how much does non-randomness (selection) contribute and how much does randomness (drift) contribute? [4]
Circa 1951, Dobzhansky rejected genetic drift as a compelling explanation for variation. The motivation behind his near-reversal is not exactly clear. Empirical evidence does not seem to have been its trigger. In the 1940s and 50s, he was out-gunned by Fisher (and others) - who consigned drift to playing "a minor evolutionary role". Subsequent to his death, his intitial advocacy of drift has been - partially - vindicated. Even though drift's contribution to evolution remains equivocal, it's recognized as significant (especially on the molecular level).
After abandoning drift as an explanation for variability, Dobzhansky signed-onto an observable outcome of Mendel's first law, known as heterozygote advantage.
- Mendel's First Law
-
1. A gene can exist in more than one form.
2. Individual organisms inherit two alleles for each trait.
3. When gametes are produced by meiosis, allele pairs separate leaving each cell with a single allele for each trait.
4. After combining when the two alleles of a pair are different, one is dominant and the other is recessive.
When packed-into different genotypes, the same genes have different "fitness values". For example, the human sickle cell gene is beneficial in the heterozygous (Xx) state but deleterious (and potentially lethal) in the homozygous (XX) state. Individuals, carrying the sickle gene in the (Xx) state, are conferred an immunity against malaria, i.e. a heterozygote advantage. This consequence of Mendel's law shows how a gene (that is disadvantageous in one genotype) can spread through a population - because it is advantageous in another. As applied to long-term evolutionary trends, Dobzhansky theorized that the beneficial effects of alleles would evolve into dominance - while the deleterious effects would evolve into recessiveness.
Virtually every chromosome contains genes which are disadvantageous in the homozygous condition, yet most organisms are well-adapted to their environments because they are heterozygous for disadvantageous alleles.
"It is the adaptive level of individuals heterozygous for various chromosomes which is most important." -- Dobzhansky (1942) [5]
A later theory, his "balance view", posits that variation, occurring among organisms, is brought-about by selection preserving them on the basis of their being "highly heterozygous". Variation (after 1951) became to Dobzhansky a function of selection, not randomness - the process of selection being an active sustainer of variation. Dobzhansky's "balance view" contrasts with an influential theory, held by Muller and Fisher. Under their "classic view", selection progressively grinds variation down until only one optimal "wild-type" allele at each locus persists, and a single phenotype dominates a species.
- Epistasis
- A phenomenon whereby the effects of one gene are modified by one or more different genes
Dobzhansky pioneered the study of epistasis. For instance, one gene can mask the expression of a phenotypic trait, controlled by another gene. The classic case for epistasis is albinism (the complete lack of an individual's ability to manufacture pigment in the skin, hair and eyes due to absence or defect of an enzyme involved in the production of melanin). In albinism, a recessive allele interferes with the production of melanin on the part of another allele.
Under the theory of population genetics, such an "epistatic interdependency" among genes was downplayed - though not rejected entirely. In models of NS devised during the early Synthesis, each gene was considered to make its own unique (relatively independent) contribution to fitness against a washed-out backdrop of other genes. As accepted today, the "fitness increase" of any allele depends on many distinct alleles. Against population genetics, epistasis brought-about an understanding of NS as a "mutualistic" evolutionary driver. (The alleles - which Mayr derided as "beans" - cannot be counted Fisher-like, as individualized, atomic units - since many of them cling together, contributing interdependently to an increase or decrease in fitness.)
Dobzhansky linked epistasis to incipient aspects of speciation. The "Dobzhansky-Muller model" of speciation proposes that a postzygotic barrier to gene flow between a parent and a daughter population occurs - because of "genic incompatibility". Due to gene interactions, the consequences of reproductive isolation trigger speciation events. Epistasis at two or more loci between two diverging populations gives rise to hybrid sterility or inviability. The simplest version of this model involves interaction between only a pair of genes. As a cause of postzygotic isolation, epistasis propels the divergences of parent and daughter populations.
- Pleiotropy
- The effects of a single gene on multiple phenotypic traits
In 1927 - at the University of Leningrad using Drosophila as his experimental subjects, Dobzhansky conducted the first exhaustive investigation into pleiotropy. [6] Pleiotropy shows that there is no one-to-one correspondence between a gene (carried an organism) and a trait (expressed by that organism). Below is a visual representation of pleiotropy in action. The length of the Happy-face Hawiian spider's matching legs is controlled by the same genes, giving it a symmetrical appearance.

Pleiotropy correlated to prezygotic isolation is a major factor, pushing the widening of character differences between two populations (a parent and a daughter) undergoing speciation. Mate choice (i.e. prezygotic isolation) for the most fit individuals in the daughter population favor localized adaptations, and genes, functioning as "pleiotropic speciation genes" ("magic traits"), accelerate the genetic separation of speciating populations.
From 1958 to 1963, Dobzhansky selectively bred fruit flies until the experimental group and the control group could no longer successfully interbreed. Male offspring were sterile after a crossing between a parent of the control group and a parent of the experimental group, and the males and females of the experimental group developed a "lack of interbreeding affinity" towards those of the control group. In plain English, he artificially induced a speciation event. [7] The tendency of hybrids (especially males) to be sterile is an example of Dobzhansky's postzygotic isolation mechanisms.
Throughout his unparalleled career, Dobzhansky wrestled with the perplexing demon - which bedeviled Darwin (and every other evolutionary biologist for over 150 years): If selection pressures reduce variation, then why is there so much variation observed in nature? [8]
Notes
(1) Genetics and the Origin of Species (1937) "Since, furthermore, the only known mode of the origin of the gene variability is through mutation, it is justifiable to suppose that the variability in question arose ultimately by occurrence of mutants in nature."
(2) Regarding the nature and identity of isolating mechanisms, Dobzhansky turned-out wrong. He included features of the environment (such as mountain ranges) which restrict interbreeding between populations. Today, only factors exclusively tied to genetics are considered isolating mechanisms - proper.
(3) Gould from "Dimensions of Darwinism" [Marjorie Grene, ed 1983] pg. 89 The hardening of the modern synthesis
"Dobzhansky, the key figure in the transition, discovered that his favorite example of potential nonadaptation needed to be reinterpreted in selectionist terms. In 1937, he had attributed differences in inversion frequencies within natural populations of Drosophila to genetic drift, but he then discovered (see 1951) that these frequencies fluctuate in a regular and repeatable way from season to season, and decided (with evident justice) that they must be adaptive." -- Gould
Also, see Gould (1980).
(4) Here is a representative sample of Dobzhansky's ultimate turn-around regarding drift. An experimental study of interaction between genetic drift and natural selection (1957) Evolution: 11 pp 311-319 Dobzhansky, T. & Pavlovsky, O
"...random drift by itself is unlikely to bring about important evolutionary progress. Indeed, variations in gene frequencies induced by drift in small isolated populations are apt to be inadaptive, and hence likely to result in the extinction of such populations." -- Dobzhansky & Pavlovsky
(5) (Genetics, 27 [1942], 487)
(6) In 1910, Ludwig Plate identified the phenomenon and introduced the term pleiotropy.
"I call a unit of inheritance pleiotropic if several characteristics are dependent upon it; these characteristics will then always appear together and may thus appear correlated." -- Plate
(7) Dobzhansky, Th., and O. Pavlovsky, 1971. "An experimentally created incipient species of Drosophila", Nature 23:289-292.
(8) Theodosius Dobzhansky: A man for all seasons by FJ Ayala
- Further reading
- Collected papers from a NAS colloquium on Dobzhansky's Genetics and the Origin of Species
- One Hundred Years of Pleiotropy: A Retrospective - Frank W. Stearns
- The spandrels of San Marco and the Panglossian paradigm - Gould, S.J. & Lewontin, R.C.